|
|
| 50年をかえりみる |
生命の分子物理的背景--近代的“生命論”の始動--
和田昭允
〈相模中央化学研究所 229神奈川県相模原市西大沼4-4-1〉
約50年ほど前,東夷か南蛮かは知らないが大物理学帝国の辺境,生物科学帝国との国境地帯に「生物物理」という種族が興ってきた.この知的好奇心旺盛な狩猟民族は未開の境界領域地方を開拓し,豊かな収穫をあげ,大帝国との交易を頻繁に行ってきた.しかし,大帝国の中に確固とした市民権を獲得したとは現在でもまだ云い難いように思われる.わが国の数ある物理学教室の中にある生物物理学講座の寥々たる数(全国で数講座)にもそれが現れている.
この小文は,日本物理学会の50周年記念に当たり,物理と生物の間で宙に迷っている概念を多少なりとも明確にし,次の50年の出発点にしようという大それた考えをもって書かれている.それは,今後の新しい“生命論”は物理的基盤の上にこそ展開される筈だと私が信じているためである.この目的のために,生物にはあまり馴染みのない理工学の研究者の方々を念頭に置いて,私が身のほども知らないままに自分なりに単純化した生命の分子物理的背景を説明しようと努力した.したがって目指すところはあくまでも“森”全体を描写することであり,“木”の詳細に立ち入らない点に関しては御容赦戴きたい(わが国の生物物理の歴史および個別の22分野の現状に関しては,「生物物理」発刊200号記念号1)を参照).また“生命を納得する姿勢”については独断と偏見をもって書いたので,特にこの点に関して積極的な御意見,御批判を戴ければ幸いである.
以下の話の筋道:“生命とは一体何だろうか?”を考えることから初まり,“いま,われわれは生命をどのように納得しているか”を述べ,最後にこの分野における“物理の先覚者の足跡”を辿る.
1. 生物⊂物質,生命活動⊂物性
太陽系の他の惑星とは全く異なり,現在の地球では“非生命の秩序”と“生命の秩序”の二つを見ることができる.
前者は物理学の法則に則って作られている“素直な秩序”であり,結晶の構造を始め,物質の示す諸性質,流体の作る複雑なパターンなどに到るまで,生物に由来しない諸現象のすべてに見られるものである.フラクタル次元の物理や系のカオス的発展の問題などになると考え込まざるをえないが,後の議論の都合上一応“素直”としておく.
いま一つが生命の秩序だが,これとても物理法則の厳然たる支配下にあることは言うまでもない.それどころか生体の巨大分子(タンパク質,核酸など),膜構造,あるいは細胞,さらに筋肉のような巨視的な組織,体液の流動などのすべてが分子物理学,流体力学,電磁気学などの教科書を“あらかじめ勉強したかのように”巧妙に作られ,機能しているのに驚かされる.ただ一寸違って“素直でない”のは,一見しただけでは可能と思われる多くの状態や反応の選択肢のうちの少数(多くの場合一つ)が「遺伝情報」と呼ばれる指令によって厳密に選択され,そこで選ばれた選択肢群の連鎖は結果的に一つの回路を作り,それらの回路が集まって“分子機械”と呼んでよいようなシステムを実現していることである.例えば眼が光を受けて神経に電気パルスを送り込む回路には,フォトンを受け取るタンパク質,そのタンパク質の構造変化を認識するタンパク質,その認識を増幅するタンパク質,それを受けて神経を刺激して電気信号を発信するタンパク質などの連鎖(信号伝達回路)がある.そこには信号の(S/N)比を高める巧妙なメカニズムさえも採用されている.これらは,ある種の“複製するシステム”が子孫を作り,世代に亙って生命活動を持続するための指令であり回路である.システムがあるから指令があるのか,その逆かの議論は“卵と鶏”の関係にあるので立ち入らない.とにかく,その結果としてできた一見不可思議な“システム”がいまや地球上の至る所で幅を利かせており,これらを我々は“生物”と呼び,それらが示す諸物性を“生命活動”と呼ぶ.付け加えれば,この生命による秩序形成の分枝として我々人類が謳歌する機械文明の秩序が現れたことになる.ただその形成のメカニズムは,生物のそれが“適者生存”というtautologyで表現される,自然環境まかせの選択の結果であるのに対して,人類の場合は過去の経験を記録し解析した上で計画的に適者を作るという点で大きく違う.
以下に,生命活動の指令を作ったのは何か? その実行のための回路の実体は何か?を考えてみる.
回路ができる,つまり一連の分子素子の各々がそれぞれ複数の反応選択肢の中から目的指向的に一つを選ぶようになるのは,素子である巨大分子(主としてタンパク質,酵素)に高度の分子構造識別能力が付与されているためである.これは反応(あるいは形態形成のための相互作用)特異性と呼ばれ,親から子へ唯一つ受けつがれる分子であるDNAの構造から出発して,多くの反応の連鎖の結果作られる構造に由来する.相手を鋭く選ぶこの能力はしばしば“鍵と鍵穴”の関係に譬えられるが,この場合は“柔らかい鍵(鍵穴)”であり,ある場合には“多状態の鍵(鍵穴)”が作られている.つまり相互作用する両分子のvanderWaals表面の幾何学的な形状に加えて,分子近傍の静電ポテンシャル面の形,水素結合のドナー・アクセプターの配置,あるいは水との親和性の強弱のパターン,さらには分子構造の異性化等も含めてのポテンシャル地形の合致によって初めて“錠前が開く(反応する)”のである.巨大分子のこのような“柔らかさ”の詳細は原子集団のNewton方程式を力ずくで解く「タンパク質動力学」が記述してくれる.2)なお構造の異性化という多状態性は,反応経路のスイッチングのために積極的に使われている場合が多い.
-人間には二つの型があって,生命の機械論が実証された時代がもし来たと仮定して,それで生命の神秘が消えたと思う人と,物質の神秘が増したと考える人とがある.そして科学の仕上げ仕事は前者の人によってもできるであろうが,本当に新しい科学の分野を開く人は後者の型ではなかろうか:中谷宇吉郎3)-
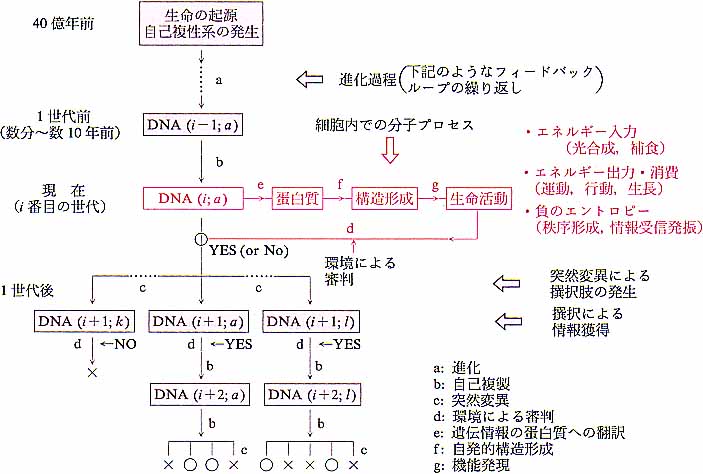
|
| 図1 遺伝情報の時間軸上,および細胞内の伝達経路.一つの分子システムが,40億年前から世代毎に環境情報をキャッチするアンテナを出し続けながら変化してきたと見られる.○:生存 ×:死滅 |
2. 生命の時間・空間・複雑さ
“素直な物性”の研究でもヒステリシスなどといって試料の過去を問題にする.
でも研究者はその現象を一つの物性と見て特に違和感を持たないだろう.しかし生物を見た場合,それも構造が最も簡単なウイルスであっても,これが物理過程の産物だとすぐ納得できるものではない.40億年と実験室的時間の圧倒的な差(十数桁)からくる違いである.
この違いの威力を納得するには,先ずとにかく生物が図1に示したような歴史を背景とした産物であることを知って戴きたい.その上で図2にイラストしたように[時間][空間][複雑さ]の3次元空間において広範な領域を占める生命の規模を見て下さい.4)この図で“素直な秩序”の殆どは時間軸近くにある一方で,宇宙や地球規模の事象はマクロ領域で生命の領域と重なるところが多い.しかし生命の特徴は,この広範なマクロ・ミクロの全領域にわたって,非常に長い緩和距離・緩和時間を持つ因果のネットワークが張り巡らされていることである.つまり何処かに刺激を与えるとその影響は広い範囲に及び,長く継続する.とにかくエイズウイルス1匹が持つDNAの1個の塩基が変わると,その影響が全世界に及び,歴史を変えるのだから.40億年の時間と全地球規模の空間が作る“超”カオスである.ここで付言すれば,現在人類社会に問われている環境問題もつきつめれば生命の持つこの特異な長距離・長時間相関にその根元がある.特に人類は,通信,大量輸送手段,記録手段と教育システムの発明によって,この時・空の相関領域と緩和時間を飛躍的に拡大したことを自覚すべきだろう.
ことの発端は約40億年前に地球上に自己複製(壊れる前に自分のコピーを作成)する分子系が出現したことである.その具体的な形は想像するほかないが,不安定でミスコピーによるいろいろな形を作りながら当時の地球の環境に適合したものが結果として残されてきた.原始地球の過酷な条件下にあって,この自己複製する分子系が不安定さを保ちながら生き延びるという二律背反の課題を克服したことはすばらしいことだったと思う.これもカオスの興味ある例題だ.複製による種族維持の連鎖がいつ切れるかも知れないという危機に曝されながらも,複製系の不安定性を安定に保持するというこの性質は失われることなく次第に洗練されて行った.その結果として現在の生物は有限の突然変異率を持つものとなった.つまり,主として複製の際にDNA分子の塩基配列に多少のランダム変更がおこり,機能タンパク質の機能が変わり,回路が変わる.そこで生じる複数の変異種は図1に示したように環境によって価値(生かすべきか死ぬべきか)が判定され,指令は結果として書き換えられて行く.このように40億年にわたる試行錯誤の結果を記憶し続けている生物が,生存のための必然として物質,エネルギー,情報の開放系,非定常系となったことは自明と思えるし,また階層構造をもった“超”複雑系とならざるを得なかったことは充分理解できる.5)ここに,従来の物理にとって全くつきあい難いものが出来上がってしまったのである.
この系の進化の過程における環境情報の取り込みは次のように理解できる.
ある世代に作られた複数の変異種の一部だけが環境条件によって次の世代として選ばれる(淘汰)ことは,この世代交代において,系が環境の情報を取り入れたことと等価である.
すなわち,現在の生物がもつ遺伝情報は,DNAの鎖状分子骨格上にA, T, G, Cと略記される原子団の1次元配列として書かれている.この分子文章は4文字コードだから文字当り2ビットの情報を持つ.高等生物のDNAは109ほどの文字を持つ(ヒトDNAは3×109)から,その情報量の上限は109ビット程度である.実際の遺伝情報のコードにはかなりの冗長度があり,本当の情報量はこれよりずっと少ない.
ここで生命の誕生以来の時間は109~1010年の程度であることを考えると,原始複製系から現在の生物にいたる進化の過程での遺伝情報の蓄積速度はせいぜい1ビット/年ということになる.すなわち年平均で2種類の変異を作り,その内の一つが選ばれ(1ビットの定義)れば充分ということになり辻妻が合う.
ちなみに現存する微生物の世代交代時間は1時間のオーダーであり,年間数千~数万の世代交代を行う.生命の起源以来の下等生物の世代数は数兆世代になるだろう.
このような気の遠くなるような時間経過を通して,生物の各世代は遺伝情報に変化を起こさせる「突然変異の圧力」と,変化した種を発展あるいは消滅させる「淘汰の圧力」に翻弄されながら今日に至った訳である.
この進化の駆動原理の問題を,地球上にある生物進化の歴史を解明し説明するという生物学の立場ではなく,分子物理学的な立場に立って実験的に解明しようとする試みがなされている.環境条件制御が可能な「進化リアクター」と呼ばれる微生物培養実験系に,世代交代周期が極端に速いウイルスを投入して超高速進化を実現させる研究である.6)そこでは上記の2種の圧力の精密制御が可能であり,得られた結果は定量的かつ厳密な議論に耐えうるものである.系を単純化した上で時間を圧縮したこの人工の自然を介して進化の過程を徹底的に尋問することで,遺伝情報空間(塩基配列空間)における適応度の地形を測量することができる.さらに配列空間における分子機能の地形,特異的結合能の地形,また3次元構造形成能の地形などを求めることができる.
上記の圧力を受けながら増殖する生物集団の振舞を数理科学として扱うのが,物理学での統計力学に相当する「集団遺伝学」である.7)
ここで生命の歴史における遺伝情報の“縦の伝達”の話を終え,つぎに個々の生物(細胞)にインプットされた遺伝情報の“横の伝達”経路(図1参照)を辿る生命の諸相を見ることにする.図にみられるように“縦”と“横”は,遺伝情報と環境情報のフィードバック回路で結ばれている.
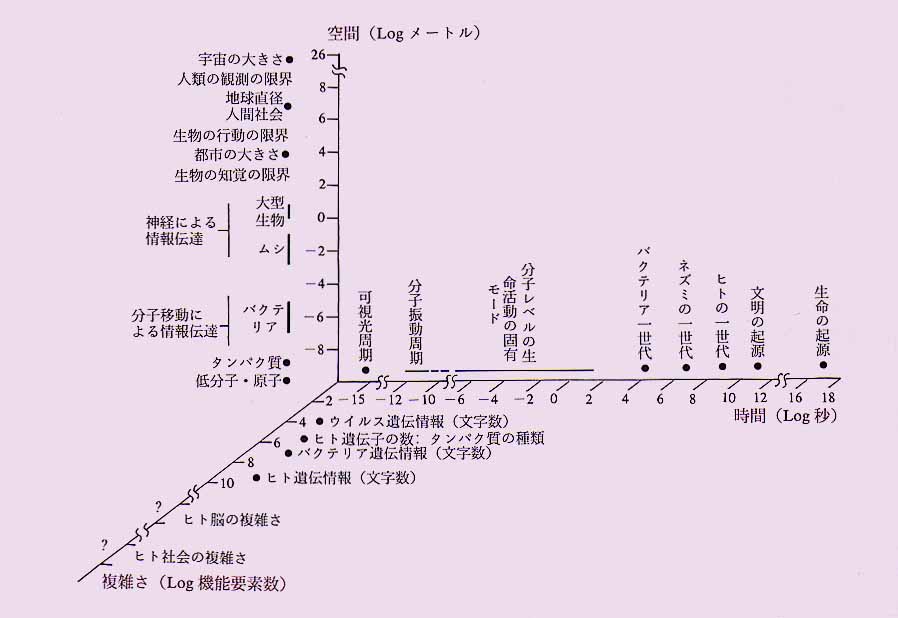
|
| 図2 生命の時間・空間領域,および複雑さ.”複雑さ”については厳密な定義はできないので,その系が持っている要素(文字,最小部品など)の概数を示してある. |
3. 生命研究の機械論的アナロジー
今日,自然科学分野の大半が生命研究に関係している.しかし,その関わり方や興味の対象が以下に述べるように大きく違うことが生命研究の奥行きの深さを物語っている.
生物は多くの分子物性をシステム化した分子機械であるという立場に立って,生命の納得の仕方を筆者の興味の赴くままに電気回路との類推によって説明してみよう.そこでは,永年物理教室にいて“物理の中華思想”のマインドコントロールから醒めきっていない私としては,分野によって理解の到達度の格段の相違(生物物理が一番理解度が高い!)を強調してしまうことになる.8,9)
もし目の前に多くの電気機器があったとすると,我々はそれらをどのように観察・研究するだろうか.まず外観をスケッチして機器の間の類似点・相違点を克明に記録・整理したり,操作用のツマミなどを動かして機械の応答を見ることから始めるだろう.「SONY」とか「NEC]等の模様の違いは,その機能とあまり本質的な関係を持たない(あるいは持つ?)ことに気づくかも知れない.いずれにせよ,これによって形態の分類表,操作に対する応答の対照表ができる.
生物学でいう「分類学」の仕事である.そこでは研究者心理の常として稀種の発見に重点がおかれることになる.生物種と物質に共通する一般法則の解明,すなわち“物理”の出動は,20世紀後半のDNA分子の遺伝情報解読の時代まで持ち越される.しかし,いまやロボット計測機器を駆使したDNA大量解析が実現し,10)遺伝情報を分子構造として比較検討することが可能になったことにより,「生物分類学」は厳密な客観性と定量性を持って再出発した.多様性の探求と共通性の解明は生物研究を支えて欠くことのできない2本柱である.
分類に次ぐ研究段階では構成要素への分解(解剖)によって構造が詳しく調べられ,大部品,中部品,小部品などが区分けされ,それらの位置関係が記述されるだろう.生物学であれば器官,組織,細胞,などの階層構造が明らかになる過程であり「形態学」と呼ばれる.聴覚の音響生理学においてHelmholtzが行ったような,器官の形態が持つ“物理”の意味づけがこの辺から顔を出し始める.
上記二つの記述的な研究に見られる因果の糸を辿る研究の始まりとして,部品間,階層間を結びつける回路の探求とそれを流れる実体の解明となる.生物学における「生理学」の分野に対応する.
機械でも生物でも回路を流れるものは「エネルギー」「物質」及び「情報」である.電気機器ではエネルギーも情報も電子によって運ばれるように設計されているが,生物の場合は分子,イオン,電子が場合によって使い分けられている点が特徴である.これは生命の回路ではミクロ化の要請によって,機械的な隔壁であるパイプや導線の使用が敬遠されたからに違いない.ただし膜構造は,細胞表面や細胞内部で隔壁であると同時に,タンパク質が機能する場としての役割を果たしている.またマクロレベルでは必要に応じて神経や血管などの導管が使われている.その代わりとして生物は,前に述べた反応特異性を活用することにより,物理的な仕切りにかえて,化学的な絶縁メカニズムによって実質的な結線を作るという設計戦略をとった.したがって,特異的相互作用のためのラベル構造と機能を装備するために,タンパク質などの生体機能分子は特徴的な構造(Schr![]() dingerの云う“繰り返しのない秩序”)を持った巨大分子とならざるを得なかったわけである.またエネルギー担体(ATPなど)は高エネルギーの燐酸結合に加えて,正しい相手にエネルギーを渡すための特異的相互作用の目印となるラベル原子団を備えたのである.
dingerの云う“繰り返しのない秩序”)を持った巨大分子とならざるを得なかったわけである.またエネルギー担体(ATPなど)は高エネルギーの燐酸結合に加えて,正しい相手にエネルギーを渡すための特異的相互作用の目印となるラベル原子団を備えたのである.
上記の3種の流れは生物学でそれぞれ「エネルギー代謝」「物質代謝」「情報伝達」と呼ばれる.これらの流れについての回路図が描かれ,新しい回路素子(タンパク質,核酸,機能膜,機能分子など)の発見が行われる.これが「生物化学」「分子生物学」の世界である.回路を辿る手段として,特定の素子を刺激(例えば特定の酵素を不活性化)して回路の作動の変化を見る,あるいはパルス信号を入れたり,流れをモジュレートして応答を見ることが行われるのは電気回路でも生物回路でも同じである.
生物の回路は複雑であり,その素子も多様(素子は生物種によっても少しずつ異なるため,その数は無限と言ってもよい;分子分類学が成り立つ由縁)であるので,まだほんの一部しか明らかになっていない.新しい機能の解明,新しい機能分子とそれが属する回路の発見がこの分野の研究者の目的なのだが,素粒子の発見と違って物理学者の興味をあまりかきたてないのはなぜだろうか? 多分,生命の“素粒子”とするには見かけ上の種類が多すぎることがひとつの理由,いまひとつは内部構造が複雑で“素”とは思えない上に,それが単に“原子が作っている積木細工”としか見えなかったからなのだろう.11)でもそろそろ化学的な複雑さの基礎に物理的な“素構造”,“素機能”が見つかりだしているのだ.
これを受けて“素子が発見され回路図が判っただけで生命活動が解明されたなどと思ってもらっては困る.それは生命理解の必要条件だが充分条件ではない”と主張する一群の研究者が登場する.それが筆者も含めた「分子生物物理」の研究者である.彼らは素子の動作特性,動作原理が“物性”として明らかになり,それを基にして回路のダイナミックスが解明され,更に生物界においてそれが採用された理由が判って初めて「なるほど,生命とはそういうものだったのか」と納得する人達である.
生命には巨視的機器では見られなかった極限のメカニズムが使われている.たとえば,熱擾乱そこそこのエネルギー(k×T(室温))入力で如何に正確に信号を伝えるか,機械を動かすか,という工夫である.生命は,分子機械の天敵である熱雑音対策を何らかの形で確立しているらしい.11)そこで我々は“如何にして(how)動作するか”の解明にとどまらず,“なぜ(why)その機構が採用されているのか”まで立ち入って理解しようとする.ここに初めて物質界に生命が存在する本当の理由が納得でき,更にそこから発展して生命の基本原理に基づくエンジニアリングがはじまる.私は,工学になるための必要条件である「デザイン」が不完全である今日,本当の「生体分子工学」はまだ始まっていないと思っている.上記のwhyの探求は価値観を伴うので物理の対象ではないかもしれないが,「生物工学」においてデザイン原理を確立するための必要条件であると思う.
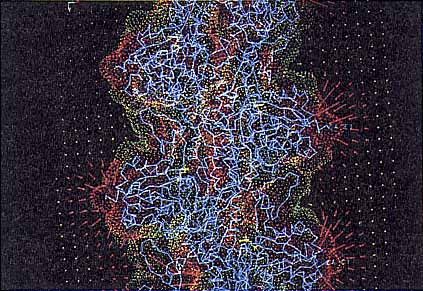
|
| 図3 アクチンフィラメントが作る電位と電場.電位は分子表面の色(青:+,赤:-)で示し,電場の大きさと方向は矢印で示してある(詳細は本文参照). |
4. 素子の研究
生物の化学回路の素子であるタンパク質,核酸,生体膜の研究のステップは,常識的には:[興味ある生命現象に目をつける]→[主役となっている素子(巨大分子,膜構造など)の抽出]→[構造解明]→[物性測定]→[前記の物質・エネルギー・情報の回路における動作特性(生物学的機能)の解明]→[分子モデルの確立]→[分子システムとして生命現象を理解]→[生命活動における共通法則の発見]となる.ただし相手が複雑なため,ステップ(たとえば構造解明)を飛ばして先に進むことも多い.
この最初の項目には,例えば「ヘモグロビンの酸素吸着」,「酵素の特異的反応」,「DNA遺伝情報の分子認識」,「感覚器における化学受容」などの特異的相互作用と分子識別,「光合成」,「光受容」などの光との相互作用,「筋収縮」,「バクテリアの鞭毛の運動」,「原形質流動」などの運動に関係した現象,「イオンチャンネル」,「イオンポンプ」などイオン輸送・制御に関係した現象,「遺伝情報発現とその制御」,「神経」,「脳」に関係した情報処理のメカニズム,等々があげられる.12)これらはとりあえず分けて書いたが,それぞれは生命活動を演出している大きなシステムの一員であるから互いに複雑に関係し合っていることは言うまでもない.また,光受容,運動のアクチュエーター,エネルギー変換,分子認識,情報処理・制御など,それぞれの素子の中および間でも,類似の分子構造や共通したメカニズムが使われている.表面的には複雑な生命も,化学種の違いに拘らずに物理構造とメカニズムに注目して一段掘り下げると,“素構造”,“素機能”と呼べるような単純な顔が出てくる.
ここで興味ある生命現象として物理の多くの研究者が目をつけてきた筋肉収縮を一例としてあげてみよう.そこでは主役(収縮),脇役(制御)を演じる素子達はすでに明らかになった.収縮情報のシグナルの経路と,それにカップルした物質輸送とエネルギー伝達の回路もかなり判った.シグナル担体の一つはCaイオンであり,エネルギー担体は前に述べた“移動分子バッテリー”ATPであって,燐酸を1個放し(放電し)てADPになるときに放出されるエネルギーが使われることが判っている.そのエネルギー収支も納得ゆくものである.11~13)しかし筋収縮の分子物理はまだ判っていない.判っているのは筋肉タンパクの一つである“アクチン”が作る繊維“アクチンフィラメント”の上を,いま一つの筋肉タンパク質であるミオシンが滑り運動をして収縮が起こるという機械モデルまでである.
しかし分子モデルの解明に向けて主役分子の構造研究,物性研究は着々として進んでいる.例えば図3に上記の“アクチンフィラメント”の周辺の電位と電場の様子を示す.フィラメント軸方向に357.5
![]() の周期をもつ原子モデルである.これを1辺が1.43
の周期をもつ原子モデルである.これを1辺が1.43
![]() の小立方体2,025,000個のメッシュで区分し,小立方体のうちのタンパク質内部に来るものの誘電率を2,外部のものは水の値の80とし,さらにイオンの存在する外部についてはデバイの遮蔽定数を考える.アクチン自身の構成原子には化学結合から予想される部分電荷を割り振る.このモデルの軸方向に周期境界条件を導入し,差分化されたPoisson-Boltzmann方程式を解く.約200万元の連立方程式を解く作業だがスーパーコンピューターで30分程度の計算時間で収束する.14)ATPによって“活性化”されたミオシン分子はこの電場の中を滑ることになるが,その1分子の出す力と変位の関係も1分子計測によって測定されている.1個の分子を相手にしてのピコ・ニュートン,ナノ・メートル領域の実時間精密計測で,まさに物理の出番にほかならない.平均化されていない1個の分子を見るからこそ,生命の熱雑音処理のメカニズムも見えてくるわけである.11,13)
の小立方体2,025,000個のメッシュで区分し,小立方体のうちのタンパク質内部に来るものの誘電率を2,外部のものは水の値の80とし,さらにイオンの存在する外部についてはデバイの遮蔽定数を考える.アクチン自身の構成原子には化学結合から予想される部分電荷を割り振る.このモデルの軸方向に周期境界条件を導入し,差分化されたPoisson-Boltzmann方程式を解く.約200万元の連立方程式を解く作業だがスーパーコンピューターで30分程度の計算時間で収束する.14)ATPによって“活性化”されたミオシン分子はこの電場の中を滑ることになるが,その1分子の出す力と変位の関係も1分子計測によって測定されている.1個の分子を相手にしてのピコ・ニュートン,ナノ・メートル領域の実時間精密計測で,まさに物理の出番にほかならない.平均化されていない1個の分子を見るからこそ,生命の熱雑音処理のメカニズムも見えてくるわけである.11,13)
しかし収縮が,分子構造の幾何学的な変化が起こす機械的な駆動が原因なのか,あるいは巧妙な静電的相互作用による分子リニアーモーター機構によるのかは,いま一息と思えるのだが全く判っていない.また,生物化学のいたるところに出てくる“活性化”という上記の表現は,エネルギーの高い状態,あるいは反応し易くなった状態という意味で漠然と使われているだけであって,その分子内物理過程の詳細描写は今後の重要な課題である.これは古典生物学で“遺伝”という言葉がその内容を極めることなしに無造作に使われていたことと同じであり,この様な重要課題に真正面から挑戦しない限り,かつて批判的に使われた“枚挙生物学”ならぬ“枚挙生物物理学”になってしまうのではないか,と心配する次第である.
この分子間の滑りは筋収縮だけでなく,バクテリアの鞭毛の回転を駆動する分子モーター,原形質流動や細胞分裂に際しての細胞内の運動機構,あるいはDNA分子鎖上の情報読み取り機構など,生命活動における[化学エネルギー]→[運動]変換に広く共通に使われている.再び強調するが,一見目の眩むように多様な化学物質とその働きは,図1の“縦”,“横”課程の複雑な干渉の結果作られた見かけ上のもので,その基盤には多くの共通性があり,そこに生命の一般原理が潜んでいる.生物物理の研究者の希望もそこにある.最後の二つの章ではこの学問の歴史を振り返ってみよう.
5. 生命の中に物理を見ようとする努力の歴史
寺田寅彦は1923年頃の随筆に次のように書いている:『物質と生命の間に橋のかかるのはまだいつの事かわからない,…最も複雑な分子と細胞内の微粒との距離ははなはだ近そうに見える.しかしその距離は全く吾人現在の知識で想像し得られないものである.山の両側から掘って行くトンネルがだんだん互いに近づいて最後の一撃でぽこりと相通ずるような日がいつ来るか全く見当がつかない.あるいはそういう日は来ないかも知れない』.15)
自然の探求は完成した絵画を知らされていないままするジグソー・パズルのようなものである.各研究者が持っている一片の部分画は,すでに出来上がっている模様(学問体系)との整合性を検証されながらそれに組み込まれていく.
1930年代から1950年にかけて生命現象に好奇心を持つ物理の研究者は,自分達が馴染んで壮大で美しい姿をすでに見せている物理科学の絵と,整合性の検証という点で捉え所のない生物科学の絵を前にして,どうしたら両方をつなぐ部分画が得られるかを模索していた.その意味で橋を架けるのに最も有利そうだと思われていた生物側の対岸は,ひとつはこれなら低分子の延長として物理で料理できるだろうという意味で寺田も書いている細胞内の巨大分子であり,いまひとつは生命活動の中の純粋な物理過程として切り出せそうな現象(たとえば,神経における電気パルス伝播,粘性流体としての血液の性質など)だった.
前者には生化学が,後者には生理学が格好な橋頭堡と思われた.後述するように,これらの分野の人達が,後に物理の研究者と協力して生物物理学を発足させて行くことになる.
寺田寅彦が書いているトンネルは30年後に,DNAの構造を明らかにしたWatsonとCrick,タンパク質の構造を示したKendrewとPerutzによって開通する.しかしその前に,そのあるべきシナリオを示した偉大な登場人物Ervin
Schr![]() dingerがいる.彼の「生命とは何か」が大きな知的インパクトを与えたのは,生物学と物理学との中間で宙に迷っている基礎的な概念を正面から問題にし,古典物理学と量子力学,無秩序と秩序,エントロピーと生命などの問題を手際よく取り上げ,個々の細部にとらわれず,問題を大きく科学思想全体の中に位置づけ,一つの方向を示している知的構想力の大きさにある.16)
dingerがいる.彼の「生命とは何か」が大きな知的インパクトを与えたのは,生物学と物理学との中間で宙に迷っている基礎的な概念を正面から問題にし,古典物理学と量子力学,無秩序と秩序,エントロピーと生命などの問題を手際よく取り上げ,個々の細部にとらわれず,問題を大きく科学思想全体の中に位置づけ,一つの方向を示している知的構想力の大きさにある.16)
Watsonは1950年頃の混迷を次のように回想する:『当時の状態では,何か新事実が現れる見込みもなかった.タンパク質や核酸の三次元構造についての議論はたいていは根拠のない空論であった.15年以上も研究は続けられているのに,すべてとはいわないまでも,大部分はまだ漠然としたことしか判っていなかった.確信をもって打ち出されるアイデアといえば,自分達の考えが容易に反論されることのない分野で仕事をするのを好む,生粋の結晶学者達の産物であることが多かった.…生物学者達はそんなたわごとを理解するために込み入った数学を勉強することなど無意味だと思っていた.…私(James-Watson)は,はっとして目を開かれる思いだった.モーリス(Wilkins)の話を聞くまでは,遺伝子とはつかみどころのない不規則なものではなかろうかという心配があった.しかし,いま,遺伝子は結晶しうることを知った.つまり,遺伝子とは規則正しい構造を持ったものであり,(X線結晶解析という)正攻法で解決できるということだ』.17)
物理学者には生命の探求に対する三つの態度があったというHans Frauenfelderの見方を紹介する.
『かつては物理学の対象は簡単なシステム,たとえば原子核,原子,あるいは分子に限られていた.分子のような複雑なシステムは化学者の手に委ねられ,物理学の寄与といえばせいぜい良い道具を提供するということに過ぎなかった.
しかしその頃でも,物理学が単に生物学に奉仕する以上のことができると感じていた人達がいた.例えばSchr![]() dingerは,一寸生命の領域に立ち入り,ひと業績上げたうえでまた物理という安全な天国に撤退した(…made a short excusion into biology, left a mark there, and then retreated again to the safe heaven of physics). 他の,例えばDelbr
dingerは,一寸生命の領域に立ち入り,ひと業績上げたうえでまた物理という安全な天国に撤退した(…made a short excusion into biology, left a mark there, and then retreated again to the safe heaven of physics). 他の,例えばDelbr![]() ckの様な何人かは生命科学に完全に没入してしまい,物理から転向した生物学者として大きな業績を上げた.しかし何人かの物理学者は彼らの物理教室にアクティヴな物理学者として残ったまま,物理学と生物学に橋を掛けるべく努力をしたのである』.18)ちなみに,私の先輩で第3のカテゴリーに入るのは,小谷正雄,岡小天,押田勇雄,大沢文夫,斎藤信彦,右衛門佐重雄,寺本英, * 三井利夫,* 松田博嗣,* 山本三三三,今井宣久等の人達である.
ckの様な何人かは生命科学に完全に没入してしまい,物理から転向した生物学者として大きな業績を上げた.しかし何人かの物理学者は彼らの物理教室にアクティヴな物理学者として残ったまま,物理学と生物学に橋を掛けるべく努力をしたのである』.18)ちなみに,私の先輩で第3のカテゴリーに入るのは,小谷正雄,岡小天,押田勇雄,大沢文夫,斎藤信彦,右衛門佐重雄,寺本英, * 三井利夫,* 松田博嗣,* 山本三三三,今井宣久等の人達である.
生物物理学社会の初期のまとまった動きとしては,1959年にReviewsofModernPhysicsが2号(No.1&2)にわたる特集 “Biophysical Study Program" を発行したが,これがこの学問に与えた推進力は計り知れないものがあった.1961年7月には国際純粋・応用生物物理学機構(現在のIUPAB (iophysics)の前身)が創設され,ここに世界的にまとまった流れが動き出したのである.
6. わが国における生物物理の勃興
わが国では1962年12月に小谷正雄,岡小天,杉田元宜,熊谷洋らの先見性と努力によって,それまで独自の活動を続けていた医学,工学,化学,生体物理学,量子生物学,放射線生物学などのいくつかのスクールが合流して日本生物物理学会が設立された.それは生命の本質は物理の正攻法で解明できるという確信を多くの人が持ち始め,それまで別れていた流れを一つにする力として働いたからである.
生物物理の発展前史を振り返ってみると,戦争によって学問が鎖国状態に置かれていたにもかかわらず,わが国において生物物理学が世界の流れに遅れることなく発足し発展した基礎には,上記の先覚者の努力もさることながら,戦前からのわが国の学問と教育レベルの潜在的な高さがあり,前述したように,多くの活発な学問の流れがすでに出来ていたことは特筆に値する.まとまるために多少の外圧が必要であったことは認めざるを得ないが,わが国での誕生期の生物物理は決して輸入品でなく,純粋に国産品だったのである.
すなわち,戦前から物理学者の中に生命現象に関心を持つ人達がかなりあったのである.よく知られているように,寺田寅彦の自由な精神のなかに学問の人為的な垣根は存在せず,森羅万象は総て物理的解釈の可能な対象であった.その意味で,寺田スクールに生物物理の源流の一つがみられる.特に平田森三は,割れ目の研究からキリンの縞模様を論じ,雑誌「科学」誌上で生物学者岡英通と激しい論争を展開している.この論争は最後には寺田寅彦が誌上で仲裁にはいるほどのものであった.以下の寺田の文は,そうは書いてないけれども,この論争と無縁ではないだろう:『「生物のことは物理ではわからぬ」という教典的信条のために,こうゆう(物理的に面白い)研究がいつもいつも異端視されやすいのは誠に遺憾なことである.科学の進歩を妨げるものは素人の無理解ではなくて,いつでも科学者自身の科学そのものの使命と本質に対する認識の不足である.深く考えなければならない次第である』.19)歴史が繰り返されなければよいが.
別の流れは,理化学研究所の仁科芳雄を中心とするグループにみることができ,立教大学や理化学研究所を拠点とする村地孝一,玉木英彦らのわが国の放射線生物学はそこで誕生した.
生物物理という名称の講義が行われた初期の例としては,上田蚕糸専門学校の上田秀雄のものがあり,「生物物理要論」(培風館)として1931年に出版されている.この様に戦前・戦中の歴史に埋もれた先覚の努力はまだまだ数多くあると思われる.
戦後の生物物理の発展は,それまで高分子物理学,統計力学,あるいは物理化学の研究者であった人達の努力に負うところが多い.
たとえば,小林理学研究所を中心とする杉田元宜,岡小天,押田勇雄,斎藤信彦,深田栄一らは,杉田が生物システム論,岡と深田がそれぞれ血液流動と骨の圧電性などの巨視的生体物性,押田が生体エネルギー論,斎藤が生体高分子物性と幅広くかつバランスのよいスクールを形成していた.
九州大学農学部の大森恭輔らのグループは生命現象に対する情報論的なアプローチを行ったという点で,上述の杉田と共にユニークな存在であった.
東京大学の水島三一郎スクール,あるいは大阪大学の赤堀四郎スクールからは生物物理学,分子生物学の分野で1960年以降に活躍する人達が輩出している.
これら生命に好奇の目を向けた先人達の築いた基盤の上に,現在多くの研究者達が活発な研究を展開しているが,それらについての真の評価は物理学会100周年記念号で明らかになるはずである.
7. 分子生物物理の使命--まとめに代えて
分子生物物理は生命活動の物質的基礎を解明しようとしており,物性物理の部分空間でもあるし,情報科学,さらに“複雑性の物理”の部分空間でもある.そして,本家の物理学と他の応用学問の関係と同じく,生命科学の全ての分野に基本原理,基本法則を提供する基盤科学である.と同時に,今までの物理が直面したことのない“素直でない系”,すなわち非線形・非平衡系で自己複製と自己組織力をもつ高度複雑系としての生命を相手とする地の利を生かして,その中にかいま見えてくる未発見の過程を物理学の全ての分野に分かりやすく紹介する窓口となりつつある.さらに応用面では40億年の進化過程で開発・蓄積された自然の工学原理を明らかにし,既存の工学と融合させつつ次の時代の技術的基盤を建設する希望を持っている.
そして何よりも,“宇宙論”の基盤に宇宙物理学があるように,“生命論”の基盤には生物物理学がある.その将来の発展はこの学問が“新しい生命論”の牽引車になれるか否かにかかっている.
参考文献
- 生物物理 35 (1995) No. 4.
- S. Hayward and N. Go: Annu. Rev. Phys. Chem. 46 (1995) 223-Collective variable description of native protein dynamics.
- 中谷宇吉郎:『中谷宇吉郎随筆集』岩波文庫(岩波書店,1988)-簪を刺した蛇.
- A. Wada: Nature 373 (1995) xxxix-A space-time slide rule.
- 美宅成樹:日本物理学会誌50 (1995) 255-階層構造なしに生物を理解することはできない.
- 伏見 譲:科学61 (1991) 333-実験室における分子進化.
- 松田博嗣:日本物理学会誌 37 (1982) 368-集団生物学研究のすすめ-数理生物学と統計物理学の共通分野として-.
- A. Wada: CHEMTRACTS Biochem. & Mol. Biol. 5 (1994) 61-On the molecular origin of photoreceptor noise.
- 和田昭允:日本物理学会誌47 (1992) 231-生物物理の“物理度”.
- A. Wada: Nature 325 (1987) 771-Automated high-speed DNA sequencing.
- 大沢文夫:日本物理学会誌 49 (1994) 924-生物物理:Schr
 dinger後50年.
dinger後50年. - 生物物理学会編:『生物物理学の最前線』ブルーバックス(講談社,1990).
- 柳田俊雄:電気学会誌115 (1995) 281-生物分子モーターの動作原理.
- 高橋卓也,中村春木:蛋白質・核酸・酵素 39 (1994) 1059-分子認識における静電相互作用.
- 寺田寅彦:『寺田寅彦随筆集』第1巻,岩波文庫(岩波書店,1969)-春六題(1921年4月執筆).
- 横山輝雄:『現代科学論の名著』村上陽一郎編,中公新書(中央公論社,1989)-シュレーディンガー「生命とは何か」.
- J. D.Watson著,江上不二夫・中村桂子訳:『2重らせん』(タイム・ライフ社,1968).
- Hans Frauenfelder(イリノイ大学教授;原子核物理学)より筆者への手紙.
- 寺田寅彦:『寺田寅彦随筆集』第4巻,岩波文庫(岩波書店,1969)-物質としてみた動物群(1938).
* 途中で第2のカテゴリーへと出奔.